Úvod do rozmnožovania rastlín
Rozmnožovanie rastlín môže byť nepohlavné (vegetatívne) alebo pohlavné (generatívne). Pri nepohlavnom rozmnožovaní vzniká nová rastlina z časti tela materskej rastliny, pričom vznikajú geneticky identické klony. Pri pohlavnom rozmnožovaní sa nová rastlina vyvinie zo zygoty, ktorá vznikla splynutím dvoch gamét, čo zabezpečuje genetickú variabilitu.
Základy pohlavného rozmnožovania
Pohlavné rozmnožovanie (iné názvy: pohlavné množenie, sexuálne rozmnožovanie/množenie, generatívne rozmnožovanie/množenie, sexuálna reprodukcia, generatívna reprodukcia, amfigónia) je proces, pri ktorom vzniká dcérsky organizmus, ktorý zvyčajne nesie kombináciu genetického materiálu dvoch iných (rodičovských) organizmov. V širšom zmysle zahŕňa termín pohlavné rozmnožovanie každý typ rozmnožovania, pri ktorom sa nový jedinec vyvíja z gamét (teda nie zo somatických buniek), a to aj v prípadoch, že nedošlo k oplodneniu.
Pri pohlavnom rozmnožovaní väčšinou vzniká organizmus s novou, unikátnou genetickou výbavou, ktorá nikdy nie je úplne totožná s genetickou výbavou rodičov. To je možné dosiahnuť buď výmenou génov medzi dvoma organizmami, alebo (častejšie) spojením buniek dvoch organizmov, z ktorých každý nesie svoje gény.
Gaméty a zygota
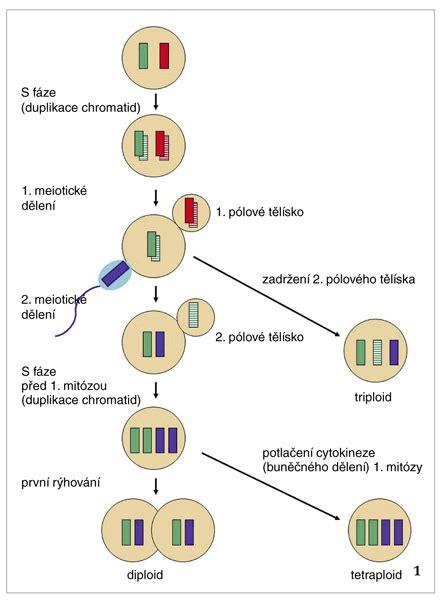
V prípade najbežnejšieho typu pohlavného rozmnožovania, eugamie, je nevyhnutné splynutie dvoch špecializovaných buniek - gamét. Ich splynutím vzniká zygota, oplodnené vajíčko, ktoré sa ďalej mitoticky delí a stáva sa z neho embryo. Gaméty zvyčajne nesú polovičnú (haploidnú) sadu chromozómov. Splynutím dvoch gamét vzniká opäť úplný (diploidný) chromozómový súbor. Ak by sa spojili dve bunky s najbežnejšou, diploidnou sadou chromozómov, vzniknutý jedinec by bol polyploidný, tzv. tetraploidný, čiže by mal až 4 súbory chromozómov. Preto je dôležité, aby spájajúce sa bunky mali len haploidnú sadu chromozómov a ich spojením by vznikol diploidný jedinec.
Genetický základ a meióza
Vlastnosti každého živého organizmu sú určené jeho deoxyribonukleovou kyselinou - DNA. Určité úseky DNA sa nazývajú gény. DNA v jadre eukaryotov je rozdelená do samostatných, rôzne dlhých úsekov, ktoré sú v spojení s bielkovinami formované do hmoty nazývanej chromatín. V čase delenia bunky sa chromatín mení na zreteľne odlíšiteľné útvary - chromozómy. Počet chromozómov je u rôznych organizmov rôzny.
Keďže pri bežnom delení buniek je väčšinou genetická výbava materských a dcérskych buniek rovnaká, pohlavné bunky vznikajú iným spôsobom ako telové (somatické) bunky, a to redukčným delením - meiózou. Meióza je zvláštnym typom delenia, pri ktorej namiesto dvoch dcérskych buniek vznikajú až 4 dcérske bunky, ale každá len s haploidným počtom chromozómov. Meióza je základným spôsobom vzniku pohlavných buniek.
Pohlavie a jeho význam
Nie každá haploidná bunka sa môže spojiť s každou. Možné je spojenie len dvoch buniek rozdielneho pohlavia. Pohlavie je vlastnosť zabraňujúca spojiť sa gaméte s gamétou, ktorá má rovnakú túto vlastnosť. Väčšinou je pohlavie dané geneticky, no k jeho vývoju či dokonca zmene môže dôjsť až v priebehu života. Ak je jedinec schopný tvoriť samčie aj samičie pohlavné bunky, hovoríme o obojpohlavnosti (hermafrodizme). Ak jedince produkujú iba samčie alebo iba samičie pohlavné bunky, nazývajú sa gonochoristi.
Okrem pohlavia existujú mechanizmy na molekulárnej úrovni, ktoré bránia kríženiu príliš odlišných jedincov, čiže jedincov patriacich do nepríbuzných taxónov. Jedným z najjednoduchších mechanizmov je rôzny počet chromozómov v pohlavných bunkách.
Výhody a nevýhody pohlavného rozmnožovania
Výhodou pohlavného rozmnožovania je rôznorodosť vzniknutého potomstva, ktorá je dôležitá z dvoch dôvodov: umožňuje jedincom rovnakého druhu rozdiferencovať svoje životné nároky v rôznorodom prostredí a spôsobuje, že jednotliví jedinci populácie reagujú na rovnaké faktory prostredia odlišne. Ďalšou výhodou je diploidný stav genómu, kde prípadnú mutáciu jedného génu môže kompenzovať jeho druhá funkčná kópia.
Nevýhodou pohlavného rozmnožovania je jeho relatívna pomalosť v porovnaní s nepohlavným. Jedince sa môžu pohlavne rozmnožovať až po dosiahnutí pohlavnej dospelosti, a často tiež vyprodukuje menší počet jedincov. Ďalšími nevýhodami sú zrieďovanie vlastného genetického materiálu a nutnosť zložitejšieho aparátu.
Pohlavné rozmnožovanie semenných rastlín
Rastliny sú vo veľkej väčšine prípadov schopné pohlavného rozmnožovania. U mnohých nižších rastlín, machorastov, rias a húb je typické striedanie pohlavnej a nepohlavnej generácie, tzv. rodozmena. Za rodozmenu možno považovať aj pohlavné rozmnožovanie cievnatých rastlín, pričom fáza nepohlavnej generácie (gametofytu) je u nich silne potlačená a gametofyt, ktorý bol u machorastov tvorený miliónmi buniek, ich u semenných rastlín obsahuje iba niekoľko málo. Samičia pohlavná bunka u rastlín sa nazýva oosféra, samčie bunky sú spermatozoidy alebo spermie. Suchozemské semenné rastliny sú odkázané na prenos samčích pohlavných buniek k samičím vetrom alebo živočíchmi. Semenné rastliny majú vajíčko mnohobunkové a predstavuje v podstate samičí gametofyt. Samotná samičia generatívna bunka oosféra sa vytvára až vnútri vajíčka. Pokiaľ je rastlina schopná produkovať samičie aj samčie pohlavné bunky, nazýva sa jednodomá.
Opelenie a oplodnenie
Opelenie (pollinatio) je prenesenie peľu z tyčinky na bliznu piestika. Ak je peľ prenesený z tyčinky na piestik toho istého kvetu, jedná sa o samoopelenie (autogamia). Ak je peľ prenesený na cudzí kvet, je to cudzoopelenie (allogamia). Špecifickým prípadom samoopelenia je Kryptogamia (skrytoopelivosť), pri ktorej dochádza k opeleniu ešte v púčiku. Peľ môže byť prenesený pomocou vetra (vetroopelivé rastliny), prostredníctvom živočíchov (hmyz, vtáky, malé cicavce) alebo vodou.
Vývin samčieho gametofytu (peľového zrna)
Peľové zrná vznikajú v peľnici po redukčnom delení. Vzniknuté bunky majú haploidné jadro, ktoré sa mitoticky rozdelí a vzniknú dve haploidné jadrá: vegetatívne (vyživovacie) a generatívne (rozmnožovacie). Neskôr sa generatívne jadro ešte raz rozdelí a vznikajú dve spermatické jadrá. Práve tieto spermatické jadrá sa zúčastnia oplodnenia ako samčie pohlavné bunky (gaméty). Vo vnútri peľového zrna sa oddelí niekoľko drobných buniek (samčie protálium). U ihličnanov exina odstáva a vytvárajú sa vzdušné vaky, ktoré umožňujú prenos peľových zŕn na veľké vzdialenosti.
Vývin samičieho gametofytu (vajíčka)
Vajíčka vznikajú u semenných rastlín na plodolistoch. Plodolisty nahosemenných rastlín sú tvorené vretenom a šupinami, na ktorých sú uložené nekryté vajíčka. Plodolisty krytosemenných rastlín zrastajú a tvoria piestik (gynaeceum). Na každom plodoliste sa nachádza zárodočné pletivo (placenta), kde sa vytvárajú vajíčka.
Vajíčko u rastlín je mnohobunkový útvar, ktorý pokrývajú obaly - integumenty. V obaloch je otvor - mikropyla (peľový vchod), ktorý umožňuje samčím gamétam preniknúť do vajíčka a oplodniť vajcovú bunku. Vo vnútri vajíčka sa nachádza pletivo nucelus, ktorého jedna bunka - materská bunka - sa zväčšuje a redukčne sa delí. Vytvorené haploidné bunky tvoria samičiu megaspóru - nezrelý zárodočný miešok. U väčšiny rastlín tri z týchto buniek zanikajú a zostane len jedna bunka, ktorá sa ďalej vyvíja.
Vývin zárodočného mieška u nahosemenných rastlín
U nahosemenných rastlín sa jadro zostávajúcej spóry mnohonásobne mitoticky delí a vzniká veľký počet haploidných jadier. Po vytvorení bunkových priehradok vzniká zásobné pletivo - primárny endosperm. V ňom sa postupne diferencuje jeden alebo viac zárodočníkov (archegónium) s vajcovou bunkou (oosféra). Tento útvar je zrelý zárodočný miešok u nahosemenných rastlín. Archegónium je samičí pohlavný orgán rastlín, oosféra je samičia gaméta.
Vývin zárodočného mieška u krytosemenných rastlín
U krytosemenných rastlín sa zostávajúca spóra mitoticky delí 3-krát, vytvorí sa bunka s ôsmimi haploidnými jadrami. Tento útvar sa označuje ako zrelý zárodočný miešok krytosemenných rastlín.
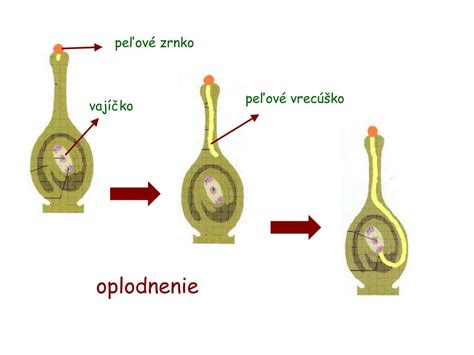
Priebeh oplodnenia
Zrelý zárodočný miešok semenných rastlín je po opelení pripravený na oplodnenie. Ak sa dostane zrelé peľové zrno na bliznu piestika, vlhká blizna spôsobí, že peľové zrno začne klíčiť. Vytvára sa peľové vrecúško, ktoré prerastá cez čnelku až do semenníka. Do peľového vrecúška sa presúvajú vegetatívne jadro a obe už vytvorené spermatické jadrá. Peľové vrecúško prerastá až k obalom vajíčka, praská a spermatické jadrá sa dostávajú cez mikropylárny otvor dovnútra do vajíčka.
- U nahosemenných rastlín splynie spermatické jadro s vajcovou bunkou a vytvorí sa zygóta. Zásobné pletivo (endosperm) je haploidné a je vytvorené pred oplodnením.
- U krytosemenných rastlín splynie jedno spermatické jadro s vajcovou bunkou - vzniká zygóta. Druhé spermatické jadro sa spojí s centrálnym jadrom zárodočného mieška a postupne sa z neho delením vytvorí triploidný (3n) endosperm.
Splynutím samčej a samičej gaméty nastalo oplodnenie. Zo zygoty sa ďalším delením a diferenciáciou vyvíja embryo. Celé vajíčko sa premieňa na semeno. Z vnútornej vrstvy integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha rozširovaniu semien vetrom. Pre proces embryogenézy je charakteristické voľnojadrové delenie, kedy sa jadro zygoty delí bez tvorby bunkových stien. U ihličnatých rastlín je zvláštnosťou polyembryónia, kedy sa z jednej zygoty môže vyvinúť viac embryí.
Hnojenie a tvorba semien
Typy semenných rastlín
Nahosemenné rastliny (Gymnospermae)
Nahosemenné rastliny sú veľký taxón semenných rastlín, ktoré predstavujú evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Vyvinuli sa koncom starších prvohôr. Dnes majú listy zväčša ihlicovité alebo šupinovité a sú prevažne vždyzelené.
Charakteristika nahosemenných rastlín
Najdôležitejším znakom nahosemenných rastlín je tvorba semien. Vajíčka a z nich vzniknuté semená nie sú ukryté v semenníku (netvoria plody), ale ležia voľne exponované na semenných šupinách, preto sa nazývajú „nahé“. Chýbajú im typické kvety s obalmi a bliznou. Kvety sa zoskupujú do jednopohlavných samčích a samičích šišiek (strobilov). Termín „plod“ sa v botanickom zmysle vzťahuje výlučne na krytosemenné rastliny, pretože vzniká zo steny semenníka. Nahosemenné rastliny, ktoré nemajú semenník, z neho nemôžu vyvinúť plod. Opelenie u nahosemenných rastlín prebieha prevažne pomocou vetra. Vytvárajú peľovú trubicu, čím sa pri oplodnení oslobodili od vody. Peľ prechádza priamo k vajíčku a oplodnenie zabezpečuje len jedna spermatická bunka. Vodivé pletivá (xylém) sú jednoduchšie - namiesto pravých ciev obsahujú len cievice (tracheidy), čo im dáva charakter mäkkého dreva. Mnohé druhy si pletivá chránia tvorbou živice.
Rozdelenie a zástupcovia nahosemenných rastlín
V starších systémoch sa všetky recentné druhy zaraďovali do oddelenia borovicorastov (Pinophyta), ktoré malo dve hlavné vývojové vetvy: ihličnany a ginká.
- Fosílne skupiny:
- Lyginodendrorasty (Lyginodendrophyta), známe aj ako semenné paprade, sú fosílne dreviny vyskytujúce sa od mladších prvohôr až do jury.
- Kordaitorasty (Cordaitophyta) sú fosílne dreviny z karbónu, ktoré sa vyvinuli z prasličkorastov a významne sa podieľali na tvorbe ložísk čierneho uhlia.
- Žijúce skupiny:
- Cykasorasty (Cycadophyta) sú „žijúce fosílie“ s hrubým, často šupinatým kmeňom a dlhými perovito zloženými listami. Sú striktne dvojdomé a opeľuje ich hmyz.
- Ginkorasty (Ginkgophyta) predstavujú jedinečnú a starobylú vývojovú líniu, ktorú dnes reprezentuje len ginko dvojlaločné (Ginkgo biloba). Je to dvojdomá rastlina s vejárovitými listami, ktorá netvorí klasické šišky, ale mäsité semená s tvrdou kôstkou.
- Borovicorasty (Pinophyta) tvoria najpočetnejšiu a evolučne najúspešnejšiu skupinu nahosemenných rastlín (asi 550 druhov).
- Ihličnany (Pinopsida) sú evolučne veľmi staré dreviny s úzkymi ihlicovitými alebo šupinovitými listami. Zvyčajne ide o jednodomé rastliny tvoriace drevnaté šišky. Zástupcami sú borovice (Pinus), jedľa (Abies), smrek (Picea) a smrekovec (Larix).
- Cyprušovité (Cupressaceae) sú vždyzelené stromy a kry, napr. borievka (Juniperus) a tuja (Thuja).
- Tisovité (Taxaceae) zahŕňajú dvojdomé dreviny ako tis obyčajný (Taxus baccata).
- Tisovcovité (Taxodiaceae) zahŕňajú reliktné stromy ako sekvoja vždyzelená (Sequoia sempervirens).
- Lianovcorasty (Gnetophyta) sú evolučne najpokročilejšia skupina a dôležitý prechod ku krytosemenným rastlinám. Majú v dreve pravé cievy a u niektorých dochádza k dvojitému oplodneniu, napr. velvíčia podivná (Welwitschia mirabilis).
Krytosemenné rastliny (Angiospermae)
Krytosemenné rastliny sa líšia od nahosemenných tým, že ich vajíčka a následne semená sú kryté v semenníku, ktorý sa po oplodnení vyvíja do plodu. U krytosemenných rastlín sa za opelenie považuje prenos peľového zrnka na bliznu. Charakteristickým rysom je dvojité oplodnenie, pri ktorom jedno spermatické jadro oplodňuje vajcovú bunku, zatiaľ čo druhé splynie s centrálnym jadrom zárodočného mieška, čo vedie k vzniku triploidného endospermu.
Nepohlavné (vegetatívne) rozmnožovanie rastlín
Nepohlavné rozmnožovanie vyžaduje len jeden materský organizmus a prebieha výlučne mitózou, čím vznikajú geneticky identické klony. Je to energeticky výhodná stratégia na rýchlu kolonizáciu stabilného prostredia. Nahosemenné rastliny sa vegetatívne rozmnožujú len zriedka. Medzi rastlinami vo všeobecnosti existujú rôzne formy nepohlavného rozmnožovania, ako sú napríklad výtrusy (u nižších rastlín) alebo špecializované útvary vzniknuté premenou stonky, koreňa či listov (napr. odnože, hľuzy).
Rozmnožovanie odrezkami
Jednou z metód vegetatívneho rozmnožovania je zakorenenie odrezkov v substráte alebo vo vode. Na rozmnožovanie potrebujeme odrezok s dvoma pármi listov. Spodný pár listov odrežeme a vrchné listy ponecháme, prípadne ak sú väčšie, zmenšíme ich o polovicu alebo 2/3 ich pôvodnej veľkosti. Týmto zabránime nadmernému odparovaniu vody cez listy. Odrezok skrátime pod spodnými púčikmi na menej ako 1 cm a spodok odrezku zasypeme práškovým stimulátorom zakoreňovania alebo namočíme do tekutého stimulátora podľa návodu výrobcu. Potom odrezok zapichneme do vlhkého substrátu (najlepšie špeciálny substrát na zakoreňovanie) a rastlinku prikryjeme fľaškou alebo sáčkom, aby sme získali prostredie so zvýšenou vlhkosťou. Odrezok pravidelne rosíme a sem-tam vyvetráme fľašu alebo sáčok. Snažíme sa udržiavať rovnomernú teplotu (cca 22-25°C). Rastlinka by mala zakoreniť v priebehu niekoľkých týždňov.
Týmto spôsobom získavame rastliny totožné s materskou rastlinou, ktoré skoro nastupujú do rodivosti. Vo všeobecnosti je nevýhodou slabší koreňový systém a to, že rastlinka nie je aklimatizovaná prostrediu tak ako semenáčik.
Semená a ich úloha v rozmnožovaní
Generatívne rozmnožovanie pomocou semien sa využíva pri šľachtení rastlín a pre získavanie podpníkov na štepenie. Pri tomto spôsobe rozmnožovania sa vysievajú dobre vyzreté semená získané z vybraných rastlín. Vzniká populácia rastlín (potomstvo) s rôznymi vlastnosťami danými rôznym genetickým založením (genotypom), niekedy dosť nevyrovnaných, so spravidla bujným rastom, rôzne dlhou juvenilnou fázou (neskorší vstup do rodivosti), aloríznym koreňovým systémom (výrazný kolovitý koreň a slabšie bočné korene).
Kvalita osiva a materské stromy
OSIVO sú semená alebo plody kultúrnych rastlín, ktoré sú určené na výsev a majú mať najlepšie úžitkové vlastnosti. Musí spĺňať požiadavky vykonávajúcej vyhlášky k ustanoveniam zákona pre daný druh, čo sa týka čistoty, klíčivosti, prímesí iných druhov a zdravotného stavu.
Osivo ovocných rastlín pre dopestovanie generatívnych podpníkov sa získava z plodov uznaných alebo registrovaných materských semenných stromov. Tieto stromy majú zaručenú pravosť druhu a odrody, spĺňajú podmienky dané zákonom o odrodách, osive a sadbe pestovaných rastlín. Majú mať dobrú rodivosť, byť odolné proti chorobám a škodcom, bezvirózne a znášať extrémne podmienky. Pri výsadbách materských semenných stromov musí byť dodržaná izolačná vzdialenosť od stromov iného genetického založenia toho istého druhu. Plody by sa mali zberať v optimálnom stupni zrelosti, ktorý zabezpečí dobrú kvalitu osiva (klíčivosť) a ľahké vylúštenie.
Medzinárodná organizácia (International Seed Testing Association - ISTA) vydáva „Medzinárodné predpisy pre skúšanie osiva“. Zvlášť dôležité sú pravosť, čistota, klíčivosť a energia klíčivosti (podiel vyklíčených rastlín za určený čas).
Klíčenie semien
Klíčenie je prechod od latentného života embrya v semene do aktívneho života. Nastáva po prerušení ich latencie súvisiacej s odvodnením protoplazmy. Klíčenie semien začína prijatím vody do semien (nabobtnanie), potom sa zosilňuje dýchanie a stupňuje sa enzymatická aktivita. Vonkajším prejavom začiatku klíčenia semien je rast radikuly (korienka) prasknutou testou (osemením). Zvýšená rýchlosť dýchania pri klíčení semien je nevyhnutná, pretože sa pri ňom spotrebúva značné množstvo energie. Sú mobilizované rezervné látky uložené v zásobných pletivách semien (v endosperme resp. klíčnych listoch).
Teplotné optimum klíčenia je pri rastlinách z našej oblasti medzi 25-28 °C, maximum medzi 30-40 °C. Pri teplomilných rastlín je optimum medzi 30-35 °C, maximum 40 °C a viac. Teplotné minimum a optimum je pre klíčenie zvyčajne nižšie ako pre neskorší rast. V niektorých prípadoch pôsobí na klíčenie pozitívne kolísanie teplôt.
Klíčivosť je podiel zŕn alebo semien schopných vytvoriť morfologické znaky klíčenia v určenom čase za podmienok určených v STN 46 10 11. Vyjadruje sa v % semien, z ktorých sa počas skúšky klíčivosti vytvorili normálni klíčenci a pri ktorých sa predpokladá, že sa v priaznivých podmienkach vyvinú v normálne rastliny.
Dormancia semien a jej prekonanie
Pozberové dozrievanie je obdobie, kedy sú zberané semená v tzv. fyziologickej zrelosti. Pozberové dozrievanie vzniklo ako biologické prispôsobenie, ktoré umožňuje semenám prežiť v nepriaznivých podmienkach a bráni vyklíčeniu v dobe, kedy by vzídené rastlinky nemali vhodné podmienky pre ďalší vývoj a prežitie. Semená mnohých druhov drevín neklíčia po dozretí na jeseň, ani ak majú vytvorené optimálne vonkajšie podmienky. Semená drevín z určitých klimatických oblastí mierneho pásma podliehajú dormancii semien.
Príčiny dormancie:
- Nepriepustnosť pre vodu a plyny (najmä vrstva palisádového sklerechýmu v osemení).
- Prítomnosť látok brzdiacich klíčenie (inhibítorov) v dužine plodov, vnútri semena alebo v obale semena (napr. kyselina abscisová).
- Nedozreté embryo, ktoré po dozretí semien ešte nedosiahlo plnú veľkosť a musí sa ďalej vyvíjať.
- Fyzikálny kľud embrya, spôsobený inhibíciou, prekonateľný pôsobením chladu.
Metódy prekonania dormancie:
- Studené a teplé mokré ošetrenie: Semená sa miešajú s vlhkým pieskom a uložia sa pri teplote 15-20 °C (teplé mokré ošetrenie) a potom pri 3-5 °C (studené mokré ošetrenie).
- Stratifikácia: Udržiavanie osiva v priebehu zimy v podmienkach, ktoré sú optimálne pre pozberové dozrievanie a udržanie klíčivosti, prípadne podporu hromadného klíčenia po výseve odbúraním inhibítorov klíčenia. Osivo sa vrství alebo premiešava s vlhkým substrátom pri teplote 2-8 °C.
- Mechanické narušenie obalov (skarifkácia): Prekonanie tvrdej šupky osemenia, ktorá bráni prístupu vody a plynov. Semená sa môžu mechanicky narušiť (napr. v bubne so sklenenými črepinami), alebo pôsobením mikroorganizmov v pôde.
- Chemické ošetrenie: Semená sa ponoria do koncentrovanej kyseliny sírovej (podľa druhu až 2 hodiny), následne sa opláchnu. Alebo sa ošetrujú horúcou vodou (krátko alebo namáčanie na viac hodín).
- Odstránenie inhibítorov: Inhibítory klíčenia možno odstrániť vyplavením semien z drvených plodov prúdiacou vodou.
Semená za normálnych podmienok klíčivé sa môžu pôsobením nevhodných podmienok prostredia dostať do sekundárnej dormancie, napr. pri vysokých teplotách skladovania.
Uchovávanie osiva a jeho úprava
Medzi zberom a výsevom je takmer pri všetkých druhoch drevín určité časové obdobie. Osivo sa musí do sejby uchovať tak, aby sa jeho klíčivosť ostala plne zachovaná. Rozhodujúci vplyv na uchovanie osiva má jeho vlhkosť.
- Krátkodobé uskladnenie (do jedného roka): Uskladnenie vo vlhku a chlade, často s použitím stratifikácie.
- Dlhodobé uskladnenie: Slúži najmä na viacročné udržanie osiva drevín plodiacich v dlhších časových intervaloch. Možno ho použiť len pri osive, ktoré znáša výrazné zníženie vlhkosti, čo vedie k zníženému dýchaniu a nižšej spotrebe energie (často pri ihličnanoch).
Sklady na osivo musia byť čisté, suché, ľahko vetrateľné, prípadne klimatizované a vybavené dostatočným osvetlením. Izolovaná podlaha musí zabraňovať prenikaniu vlhkosti.
Metódy úpravy osiva:
- Fyzikálne metódy morenia osiva: Prevažne využívajú teploty, ktoré sú letálne pre škodlivé činitele. V obmedzenej miere sa využívajú aj niektoré druhy žiarenia.
- Chemické metódy morenia osiva: Fungicídne, insekticídne alebo kombinované. Morenie osiva je v mnohých prípadoch účinnejšie ako plošná aplikácia pesticídov.
- Inkrustácia: Technológia morenia osiva, ktorou sa aplikujú pesticídy pomocou fixačných látok. Inkrustačná hmota umožňuje farebné tenkovrstevné povliekanie semien, pričom sa ich prirodzený tvar a rozmery nemenia.
- Obaľovanie semien: Úprava osiva, pri ktorej sa semená obaľujú do špeciálnej hmoty, aby získali jednotnú veľkosť, tvar a hmotnosť. Umožňuje presný výsev a znižuje spotrebu osiva.
tags: #semenne #rastliny #rozmnozovanie