Nahosemenné rastliny (iné názvy: nahosemenné, gymnospermy; lat. Gymnospermae) sú veľký taxón semenných rastlín. Do začiatku 21. storočia sa často uvádzalo, že nahosemenné rastliny nie sú monofyletický taxón, ale boli považované za parafyletický alebo polyfyletický. V mnohých systémoch sa preto namiesto nich uvádzali len ich jednotlivé časti ako samostatné taxóny. Avšak najnovšie molekulárne poznatky naznačujú, že nahosemenné rastliny spolu s krytosemennými rastlinami tvoria dve monofyletické vetvy, čo viedlo k návratu k pôvodnej koncepcii považujúcej skupinu Gymnospermophyta za monofyletickú.
Nahosemenné rastliny predstavujú evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Vyvinuli sa koncom starších prvohôr, kedy vplyvom suchej klímy získali malé, kožovité listy s hrubou kutikulou. Dnešné druhy majú listy zväčša ihlicovité alebo šupinovité a sú prevažne vždyzelené.

Charakteristika nahosemenných rastlín
Najdôležitejším znakom nahosemenných rastlín je tvorba semien. Vajíčka a z nich vzniknuté semená nie sú ukryté v semenníku (netvoria plody), ale ležia voľne exponované na semenných šupinách, preto sa nazývajú „nahé“. Chýbajú im typické kvety s obalmi a bliznou. Kvety sa zoskupujú do jednopohlavných samčích a samičích šišiek (strobilov). Je dôležité zdôrazniť, že termín „plod“ sa v botanickom zmysle vzťahuje výlučne na krytosemenné rastliny, pretože vzniká zo steny semenníka. Nahosemenné rastliny, ktoré nemajú semenník, z neho nemôžu vyvinúť plod.
Opeľovanie u nahosemenných rastlín prebieha prevažne pomocou vetra. Vytvárajú peľovú trubicu, čím sa pri oplodnení oslobodili od vody. Peľ prechádza priamo k vajíčku a oplodnenie zabezpečuje len jedna spermatická bunka.
Špecifická je aj stavba ich dreva. Vodivé pletivá (xylém) sú jednoduchšie - namiesto pravých ciev obsahujú len cievice (tracheidy), čo im dáva charakter mäkkého dreva. Mnohé druhy si pletivá chránia tvorbou živice.
Rozdelenie a zástupcovia
V starších systémoch sa všetky recentné druhy zaraďovali do oddelenia borovicorastov (Pinophyta), ktoré malo dve hlavné vývojové vetvy: ihličnany a ginká.
Fosílne skupiny
- Lyginodendrorasty (Lyginodendrophyta), známe aj ako semenné paprade, sú fosílne dreviny vyskytujúce sa od mladších prvohôr až do jury. Dnes sa nepovažujú za ucelené oddelenie, ale za parafyletickú skupinu.
- Kordaitorasty (Cordaitophyta) sú fosílne dreviny z karbónu, ktoré sa vyvinuli z prasličkorastov a významne sa podieľali na tvorbe ložísk čierneho uhlia.
Žijúce skupiny
- Cykasorasty (Cycadophyta) sú „žijúce fosílie“, ktoré v druhohorách tvorili obrovskú časť vegetácie. Sú to stromovité formy s hrubým, často šupinatým kmeňom a dlhými perovito zloženými listami na jeho vrchole. Vytvárajú semená s nápadným mäsitým obalom (sarkotestou). Do súčasnosti sa zachovala len trieda cykasy (Cycadopsida). Sú striktne dvojdomé a opeľuje ich hmyz.
- Ginkorasty (Ginkgophyta) predstavujú jedinečnú a starobylú vývojovú líniu. Dnes prežíva už len jediný reliktný druh - ginko dvojlaločné (Ginkgo biloba). Je to dvojdomá „žijúca fosília“ s charakteristickými vejárovitými listami. Netvorí klasické šišky, ale mäsité semená s tvrdou kôstkou.
- Borovicorasty (Pinophyta) tvoria najpočetnejšiu a evolučne najúspešnejšiu skupinu nahosemenných rastlín (asi 550 druhov).
- Ihličnany (Pinopsida) sú evolučne veľmi staré dreviny, známe už od konca karbónu. Majú úzke ihlicovité alebo šupinovité listy s hlboko vnorenými prieduchmi a hrubou kutikulou. Zvyčajne ide o jednodomé rastliny tvoriace drevnaté šišky. Sú však citlivé na znečistenie prostredia.
- Čeľaď borovicovité (Pinaceae): Zástupcovia majú vo svojich pletivách živicové kanáliky. Ihličnany z rodu Pinus (borovice) majú ihlice vo zväzočkoch. Bežné sú borovica lesná (Pinus sylvestris), borovica čierna (Pinus nigra), borovica limbová (Pinus cembra) a borovica horská (kosodrevina, Pinus mugo). Významným druhom je jedľa biela (Abies alba). Z rodu smrekov je pôvodný smrek obyčajný (Picea abies). Jediným opadavým ihličnatým stromom je smrekovec opadavý (Larix decidua). Z nepôvodných druhov sa pestuje duglaska tisolistá (Pseudotsuga menziesii).
- Čeľaď cyprusovité (Cupressaceae): Vždyzelené stromy a kry rozšírené po celom svete. Listy sú v dospelosti najčastejšie šupinovité. Z domácich druhov je významná borievka obyčajná (Juniperus communis) s dužinatou šuškou (galbulus). Z nepôvodných je častá tuja západná (Thuja occidentalis).
- Čeľaď tisovité (Taxaceae): Zahŕňa dvojdomé dreviny. Kľúčovým zástupcom je tis obyčajný (Taxus baccata) s červeným sladkým mieškom obklopujúcim semeno.
- Čeľaď tisovcovité (Taxodiaceae): Zahŕňa reliktné stromy. Absolútnym výškovým rekordérom je sekvoja vždyzelená (Sequoia sempervirens).
- Lianovcorasty (Gnetophyta): Evolučne najpokročilejšia skupina a dôležitý prechod ku krytosemenným rastlinám. Majú v dreve pravé cievy a u niektorých dochádza k dvojitému oplodneniu. Extrémny zástupca je velvíčia podivná (Welwitschia mirabilis).

Rozmnožovanie nahosemenných rastlín
Rozmnožovanie rastlín môže byť nepohlavné (vegetatívne) alebo pohlavné (generatívne). Pri nepohlavnom rozmnožovaní vzniká nová rastlina z časti tela materskej rastliny, pričom vznikajú geneticky identické klony. Pri pohlavnom rozmnožovaní sa nová rastlina vyvinie zo zygoty, ktorá vznikla splynutím dvoch gamét, čo zabezpečuje genetickú variabilitu.
Pohlavné rozmnožovanie
V procese pohlavného rozmnožovania dochádza k tvorbe gamét. Samčie gaméty vznikajú v peľniciach (mikrosporangiách) a samičie gaméty (oosféra) v zárodočníkoch (archegóniách).
Vývin samičieho gametofytu (megagametogenéza)
Vajíčka nahosemenných rastlín sú krasinucelátne. Z primárneho archespóru, ktorý vzniká z jednej alebo viacerých subepidermálnych buniek nucela, sa po delení vytvorí sekundárny archespór. Z neho sa diferencujú megasporocyty.
Megasporocyt sa redukčne delí a vzniká tetráda haploidných megaspór. Pri niektorých taxónoch vznikajú len tri megaspóry. Zvyčajne sa vyvíja len jedna megaspóra, z ktorej niekoľkonásobným mitotickým delením vzniká voľnojadrové prothálium (samicí gametofyt). Výskyt veľkého počtu voľných jadier je charakteristický pre všetky nahosemenné rastliny. Neskôr sa vytvoria bunkové steny. Z týchto haploidných buniek vzniká endosperm a archegóniá, ktoré obsahujú oosféru.
Vývin samčieho gametofytu (mikrogametogenéza)
Vo vnútri juvenilného mikrosporangia (peľnice) sa nachádza peľotvorné pletivo (archespór). Redukčným delením buniek sporogénneho pletiva vzniknú tetrády haploidných mikrospór (peľových zŕn). U ihličnanov exina odstáva a vytvárajú sa vzdušné vaky, ktoré umožňujú prenos peľových zŕn na veľké vzdialenosti.
Mikrospóry sa začínajú deliť ešte v sporangiu. Vo vnútri peľového zrna sa oddelí niekoľko drobných buniek (samčie prothálium). Veľká bunka vo vnútri peľového zrna sa mitoticky delí na vegetatívnu bunku (zabezpečuje rast peľovej trubice) a generatívnu bunku. Generatívna bunka sa ďalej delí na nástennú bunku a spermatogénnu bunku, z ktorej vzniknú dve spermatické bunky (u primitívnejších skupín sa vyvíjajú ako obrvené spermatozoidy).
Výbor parlamentu pre hospodárske záležitosti za účasti ministerky Denisy Sakovej
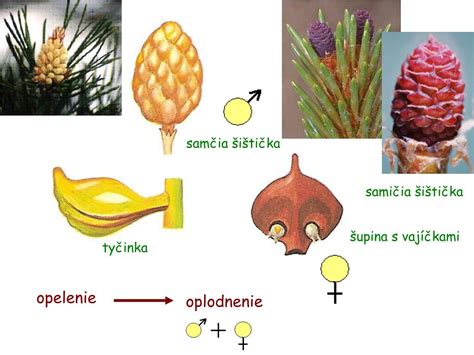
Opelenie a oplodnenie
Opelenie je prenos peľových zŕn na samičie rozmnožovacie orgány. U nahosemenných rastlín ho zabezpečuje prevažne vietor. Peľové zrná dopadajú na semenné šupiny samičích šištíc a zachytávajú sa do polinačnej kvapky vylučovanej v oblasti mikropyly vajíčka. Pri vysýchaní kvapky sa peľové zrno dostane na povrch nucela.
Peľové zrno klíči a vytvára peľové vrecúško, ktoré prerastá cez nucelus k archegóniu. Spermatogénna bunka sa v peľovom vrecúšku rozdelí na dve spermatické bunky. Jedna spermatická bunka oplodní oosféru, čím vzniká diploidná zygota. Ostatné bunky peľového vrecúška zanikajú. Oplodnená býva zvyčajne iba jedna vajcová bunka.
Vývin embrya a vznik semena
Zo zygoty sa ďalším delením a diferenciáciou vyvíja embryo. Celé vajíčko sa premieňa na semeno. Z vnútornej vrstvy integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha rozširovaniu semien vetrom.
Pre proces embryogenézy je charakteristické voľnojadrové delenie, kedy sa jadro zygoty delí bez tvorby bunkových stien. Počet voľnojadrových delení je u jednotlivých rodov rôzny. Napríklad u Ginkgo biloba nastáva cytokinéza až po vytvorení 256 jadier, u ihličnanov po vytvorení 2 až 64 jadier.
Ďalšou zvláštnosťou ihličnatých rastlín je polyembryónia, kedy sa z jednej zygoty môže vyvinúť viac embryí. Napríklad v rode Pinus sa v prvej fáze voľnojadrového delenia vytvoria jadrá, ktoré sa delia a diferencujú do vrstiev, pričom vznikajú iniciály zárodkov, z ktorých sa postupne vyvíjajú embryá.
Niektoré ihličnany, ako napríklad tis (Taxus baccata), produkujú vajíčka v mäsitých pohárikovitých arilusoch. Tis je dvojdomá rastlina.
Nepohlavné (vegetatívne) rozmnožovanie
Nepohlavné rozmnožovanie vyžaduje len jeden materský organizmus a prebieha výlučne mitózou, čím vznikajú geneticky identické klony. Je to energeticky výhodná stratégia na rýchlu kolonizáciu stabilného prostredia.
Nahosemenné rastliny sa vegetatívne rozmnožujú len zriedka. Medzi rastlinami vo všeobecnosti existujú rôzne formy nepohlavného rozmnožovania, ako napríklad:
- Výtrusy: Jednobunkové útvary na šírenie, tvorené u nižších húb a organizmov aj mitoticky.
- Špecializované útvary: U vyšších rastlín si vyvinuli špecializované orgány vzniknuté premenou stonky, koreňa či listov, napr. rozmnožovacie cibuľky, hľuzy, podzemky, poplazy (stolóny) a listy.
tags: #rozmnozovanie #nahosemennych #rastlin #ppt