Z fyziologického hľadiska rozdeľujeme spôsoby rozmnožovania rastlín do dvoch základných skupín: nepohlavné rozmnožovanie a pohlavné rozmnožovanie.
Rodozmena (Metagenéza)
Rodozmena predstavuje životný cyklus rastlín charakterizovaný striedaním dvoch geneticky odlišných generácií. Fúziou haploidných gamét (pohlavných buniek) vzniká diploidná zygota, čím sa cyklus neustále opakuje a generácie sa striedajú.
Gametofyt (pohlavná generácia)
Gametofyt tvoria bunky s jedným súborom chromozómov (haploidný počet, n). Vzniká mitotickým delením zo spóry. Neskôr vytvára pohlavné orgány (gametangiá), ktoré mitózou produkujú pohlavné bunky (gaméty): plemenníčky (anterídiá) tvoria samčie gaméty (často bičíkaté spermatozoidy vyžadujúce vodu) a zárodočníky (archegóniá) tvoria samičiu bunku (oosféra).
Sporofyt (nepohlavná generácia)
Sporofyt disponuje homologickými chromozómami (diploidný počet, 2n). Vzniká mitotickým delením zygoty. Prechod rastlín na súš znamenal evolučný tlak na ochranu citlivých fáz. Vývoj smeroval k posilneniu diploidného sporofytu a redukcii haploidného gametofytu.
Prvé suchozemské rastliny, ako ryniorasty, mali ešte primitívnejšiu rovnakotvarú rodozmenu. Výraznou evolučnou výnimkou sú machorasty (Bryophyta), u ktorých dominuje haploidný gametofyt (zelený mach). Sporofyt (stopka s výtrusnicou) je u nich fyziologicky úplne závislý od gametofytu.
U cievnatých výtrusných rastlín už dominuje mohutný diploidný sporofyt. Ich drobný gametofyt (prvorast) je síce zredukovaný, ale žije nezávislým životom. Pri semenných rastlinách je gametofyt extrémne zredukovaný a trvalo ukrytý v materskom sporofyte.
Vedeli ste, že...? Samičí gametofyt krytosemenných rastlín je zredukovaný len na 7 buniek (zárodočný vak) a samčí gametofyt dokonca len na 2-3 bunky (peľové zrnko).

Nepohlavné (vegetatívne) rozmnožovanie
Nepohlavné rozmnožovanie vyžaduje len jeden materský organizmus. Bunkové delenia prebiehajú výlučne mitózou, vďaka čomu vznikajú geneticky identické klony. Je to energeticky výhodná stratégia na rýchlu kolonizáciu stabilného prostredia bez závislosti od opeľovačov.
Nepohlavné rozmnožovanie je kľúčové pre druhy s poruchami meiózy, napríklad pri zmnožení chromozómových sád (polyploidia).
Základné fyziologické formy nepohlavného rozmnožovania
- Bunkové delenie - typické pre sinice a jednobunkové riasy.
- Fragmentácia stielky - rozpad stielky na životaschopné časti. Využívajú ju napríklad mnohobunkové sladkovodné spájavky, ktorých vlákno po roztrhnutí jednoducho dorastie na novú riasu.
- Výtrusy - jednobunkové útvary na šírenie, u nižších húb a organizmov tvorené aj mitoticky.
Špecializované orgány na nepohlavné rozmnožovanie u vyšších rastlín
Vyššie rastliny si na vegetatívne rozmnožovanie vyvinuli špecializované orgány vzniknuté premenou stonky, koreňa či listov:
- Rozmnožovacia cibuľka - pozostáva z podcibulia a zdužnatených zásobných listov. Tento typ množenia je charakteristický pre čeľaď ľaliovité (napríklad obľúbený tulipán alebo cesnak, ktorý vytvára zložené dcérske cibuľky) a skorú jarnú čeľaď amarylkovité (snežienka, narcis).
- Podzemok (rizóm) - je vodorovne rastúca podzemná stonka. Z jej uzlov rastú korene a púčiky formujúce nové rastliny. Je to bežný jav predovšetkým u papradí, z kvitnúcich rastlín ho využíva napríklad liečivý kostihoj alebo mimoriadne odolný burinný pýr.
- Podzemková hľuza - je zhrubnutý zásobný podzemok stonkového pôvodu. Najlepším príkladom je zemiak, ktorého známe „očká“ sú v skutočnosti pazušné púčiky novej rastliny.
- Stonková hľuza - je nadzemná alebo podzemná metamorfóza plniaca zásobnú aj reprodukčnú funkciu. Typickým zástupcom s nadzemnou hľuzou je kaleráb. Veľmi podobný mechanizmus, avšak vo forme koreňovej hľuzy, si vytvorila reďkovka.
- Poplaz (stolón) - ide o horizontálnu plazivú stonku, ktorá rastie po povrchu a zakoreňuje priamo vo svojich uzloch. Tento efektívny mechanizmus plošného šírenia má jahoda.
- Listy - na nepohlavné rozmnožovanie môžu slúžiť priamo aj listy.
Pohlavné rozmnožovanie
Pohlavné rozmnožovanie spája genetický materiál dvoch jedincov, čím zabezpečuje nevyhnutnú genetickú variabilitu. Nová rastlina vzniká zo zygoty, produkovanej splynutím dvoch haploidných gamét.
Samčie a samičie gametofyty
Peľové zrná (samčí gametofyt) vznikajú v peľniciach. Ich povrch chráni pevná vonkajšia exina a vnútorná tenká intina. Zrelé peľové zrnko krytosemenných rastlín tvoria tri bunky: jedna vegetatívna bunka (zabezpečuje rast peľovej trubice) a dve samčie neobrvené bunky, takzvané spermácie.
Zárodočný miešok (samičí gametofyt) sa vyvíja vo vajíčku semenníka.
Opelenie a oplodnenie
Prenos peľu na bliznu označujeme ako opelenie. Podľa pôvodu peľu rozlišujeme samoopelenie (autogamiu) a cudzoopelenie (alogamiu). Rastliny sa samoopeleniu bránia autoinkompatibilitou (biochemický blok) alebo časovým nesúladom dozrievania orgánov.
Dômyselný priestorový mechanizmus na podporu cudzoopelenia využíva prvosienka jarná. Jednotlivé rastliny tvoria buď kvety s dlhými čnelkami a krátkymi tyčinkami, alebo presne naopak, čo priamo núti hmyz prenášať peľ výlučne medzi rôznymi jedincami.
Ak na bliznu dopadne správny peľ, prítomný vápnik, bór a fytohormóny ho okamžite stimulujú ku klíčeniu.
Dvojité oplodnenie
Proces dvojitého oplodnenia je unikátom krytosemenných rastlín. Po vniknutí trubice do vajíčka prebehnú dve splynutia súčasne:
- Prvá spermácia splynie s oosférou, čím vzniká diploidná zygota (budúce embryo).
- Druhá spermácia splynie s centrálnou bunkou, čím vzniká bunka s triploidným jadrom. Z nej sa sformuje zásobný triploidný endosperm vyživujúci embryo.
Rozmnožovanie nahosemenných rastlín
Nahosemenné rastliny (Gymnospermae) tvoria evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Vyvinuli sa koncom starších prvohôr. Ich najdôležitejším znakom je tvorba semien, ktoré nie sú ukryté v semenníku, ale ležia voľne exponované na semenných šupinách. Chýbajú im typické kvety s obalmi a bliznou; kvety sa zoskupujú do jednopohlavných samčích a samičích ších (strobíl).
Opeľuje ich prevažne vietor. Vytvárajú peľovú trubicu, čím sa pri oplodnení úplne oslobodili od vody. Peľ prechádza priamo k vajíčku a oplodnenie zabezpečuje len jedna spermatická bunka. Vegetatívne sa rozmnožujú len zriedka.
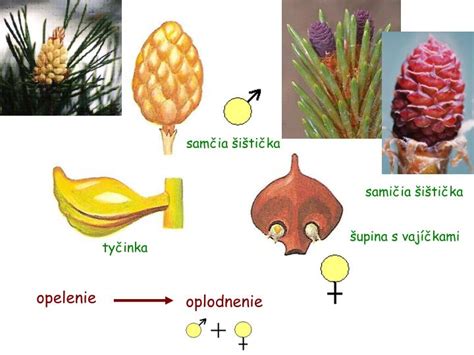
Rozmnožovanie na príklade borovice (Pinus silvestris)
Borovica, ktorá je sporofytom, má samčie a samičie šištice (strobily).
Samičia šištica (megastrobilus)
Je tvorená zdrevnateným vretenom, na ktorom sú špirálovite usporiadané podporné šupiny a v ich úžľabí vyrastajú semenné šupiny. U borovice sú podporné šupiny zakrpatené a semennými šupinami prerastené natoľko, že sú morfologicky takmer nerozoznateľné. Na vrchnej strane semenných šupín sú pri báze lokalizované dve vajíčka.
Vajíčka sa vyvíjajú z placenty tak, že sa najprv vytvorí meristematický hrbolček, ktorý je základom nucela. Z bázy vajíčka sa diferencuje kruhový val, ktorý obrastá nucelus a vytvorí sa jediný obal (integument), ktorým je vajíčko nahosemenných rastlín chránené. Na vrchole vajíčka nie je integument spojený a jeho okraje ohraničujú mikropylárny otvor.
Samčia šištica (mikrostrobilus)
Je tvorená krátkym vretenom a blanitými, špirálovite usporiadanými šupinami priliehajúcimi k vretenu. Vyrastajú v spodných etážach stromu. Blanitá šupina - mikrosporofyl - predstavuje tyčinku s dvoma alebo niekoľkými mikrosporangiami.
Vývin samičieho gametofytu
Vajíčka nahosemenných rastlín sú krasinucelátne. Nucelus je pokrytý výraznou pokožkou, ktorá je obklopená veľkým počtom parenchymatických buniek. Na vrchole je nucelus sformovaný do peľovej komory, ktorá vzniká buď tak, že skupina buniek v apikálnej časti lyzuje a vzniká plytká priehlbenina, alebo sa nápadne naddvihne a vzniká poduškovitý útvar. Oba tieto útvary slúžia na vytvorenie vhodných podmienok pre zachytenie a vyklíčenie peľového zrna.
Integument postupne prerastá nucelus a vytvorí sa mikropyla, ktorá podobne ako blizna krytosemenných rastlín, produkuje polinačnú kvapku.
Sporogénne pletivo sa zakladá z jednej alebo viacerých subepidermálnych buniek nucela. Vzniká primárny archespór. Bunky primárneho archespóru sa delia a vzniká parietálna a sporogénna vrstva. V sporogénnom pletive prebieha opakované mitotické delenie buniek, z ktorých sa niektoré transformujú na megasporocyty.
Megasporocyt sa redukčne delí a vzniká tetráda haploidných megaspór. Pri niektorých taxónoch boli pozorované iba tri megaspóry. Postavenie megaspór je zvyčajne lineárne, ale môže byť aj tetraedrické alebo v tvare T. U Pinus silvestris v procese heterotypického delenia vznikajú dve, nerovnako veľké bunky. Ďalej sa vyvíja len bazálna megaspóra, ktorej niekoľkonásobným mitotickým delením vzniká voľnojadrové prothálium. Výskyt veľkého počtu voľných jadier je charakteristickým znakom všetkých nahosemenných rastlín. V ďalšom procese sa vytvoria bunkové steny. Z týchto haploidných buniek vzniká endosperm a archegóniá. Iniciálna bunka archegónia sa delí a vzniká vonkajšia tzv. primárna hrdlová bunka a vnútorná tzv. primárna brušná bunka. Delením primárnej hrdlovej bunky vzniká hrdlo archegónia a delením primárnej brušnej bunky vzniká brušná kanálová bunka a oosféra. Samičí gametofyt Pinus silvestris teda pozostáva z haploidného endospermu a z dvoch archegónií umiestnených v oblasti mikropily. V každom archegóniu je jedna oosféra.
Endosperm má charakter zásobného parenchýmu a obsahuje predovšetkým škrob, aleuronové zrná a lipidy.
Vývin samčieho gametofytu
Samčie šištice vyrastajú spravidla na jar, na strome ostávajú niekoľko týždňov a potom opadávajú.
Vo vnútri juvenilného mikrosporangia sa nachádza peľotvorné pletivo - archespór. Z neho sa diferencuje výstelkové pletivo - tapetum a sporogénne pletivo. Redukčným delením buniek sporogénneho pletiva vzniknú tetrády haploidných mikrospór. Bunková stena vyvíjajúcich sa peľových zŕn je v tomto období tvorená exinou a vnútornou intinou. U borovice a ďalších druhov ihličnatých rastlín exina odstáva a vytvárajú sa vzdušné vaky, ktoré umožňujú prenos peľových zŕn na veľké vzdialenosti.
Pri formovaní samčieho gametofytu dochádza k primárnemu deleniu (vznik protáliových buniek), sekundárnemu deleniu (delenie anterídiovej bunky na nástennú a spermatogénnu bunku) a terciárnemu deleniu (delenie spermatogénnej bunky na dve spermatické bunky).
Mikrospóry sa začínajú deliť ešte v sporangiu. Vo vnútri peľového zrna sa v jednom mieste oddelí niekoľko drobných buniek, ktoré väčšinou po určitom čase zanikajú. Tieto bunky predstavujú samčie prothálium. Veľká bunka vo vnútri peľového zrna sa mitoticky delí. Po celularizácii vzniká väčšia vegetatívna bunka a menšia generatívna bunka. Generatívna bunka sa opäť delí a vznikne bunka nástenná a spermatogénna bunka, z ktorej pri prerastaní peľového vrecúška vzniknú 2 spermatické bunky.
U primitívnejších skupín nahosemenných rastlín (napr. cykasovitých) sa samčie gaméty vyvíjajú ako obrvené spermatozoidy.

Opelenie a oplodnenie nahosemenných rastlín
Počas prvej sezóny sa šupiny samičích šištíc so zrelými vajíčkami oddialia od vretena. Peľové zrná dopadajú medzi šupiny a na povrchu vajíčka sa zachytávajú do polinačnej kvapky, ktorá je vylučovaná v oblasti mikropily. Pri vysýchaní kvapky sa peľové zrno dostane na povrch nucela. Peľové zrno klíči, vzniká peľové vrecúško, ktoré začne prerastať cez nucelus. Šupiny samičích šištíc sa v tomto období primknú k vretenu.
Spermatogénna bunka sa v peľovom vrecúšku rozdelí na dve spermatické bunky. Jedna spermatická bunka oplodní oosféru - vzniká zygota. Ostatné bunky peľového vrecúška zanikajú. Oplodnená býva zvyčajne iba jedna vajcová bunka.
Vývin embrya a vznik semena nahosemenných rastlín
Zo zygoty sa ďalším delením a diferenciáciou vyvíja embryo a celé vajíčko sa premieňa na semeno. Z vnútornej vrstvy integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha rozširovaniu semien vetrom.
Niektoré ihličnany nemajú drevnaté šištice a nie sú ani jednodomé. Napr. tis (Taxus baccata) produkuje vajíčka na koncoch krátkych axilárnych výhonkov v mäsitých pohárikovitých arilusoch. Tis je dvojdomá rastlina.
Pre proces embryogenézy je charakteristický jav voľnojadrového delenia, to znamená, že v počiatočných fázach sa jadro zygoty delí, ale nevznikajú bunkové steny. Počet voľnojadrových delení je u jednotlivých rodov rôzny a až potom nastáva celularizácia. Napr. u Ginkgo biloba nastáva cytokinéza až po vytvorení 256 jadier, u cykasu Dioon edule 1000 a u ihličnanov po vytvorení 2 až 64 jadier.
Ďalšou zvláštnosťou ihličnatých rastlín je polyembryónia. Napr. v rode Pinus v prvej fáze voľnojadrového delenia vzniknú štyri jadrá, ktoré putujú k mikropyle a usporiadajú sa do jednej vrstvy. Jadrá sa delia, vzniká druhá vrstva jadier a súčasne sa diferencujú aj bunkové steny. Bunky sa opäť delia a vznikajú štyri vrstvy. Štyri horné - bazálne bunky, pod nimi sú ružicovité (rozetové) bunky a štyri primárne suspenzorové bunky. Na tieto sa pripájajú štyri bunky označované ako iniciály zárodkov. Z nich sa postupne vyvíjajú embryá.

Taxonomické skupiny nahosemenných rastlín
V starších systémoch sa všetky recentné druhy zaraďovali do oddelenia borovicorastov (Pinophyta), ktoré malo dve hlavné vývojové vetvy: ihličnany a ginká. Dnešné systémy považujú nahosemenné rastliny za monofyletickú skupinu.
Fosílne skupiny
- Lyginodendrorasty (Lyginodendrophyta), známe aj ako semenné paprade, sú fosílne dreviny vyskytujúce sa od mladších prvohôr až do jury. Dnes sa nepovažujú za ucelené oddelenie, ale za parafyletickú skupinu.
- Kordaitorasty (Cordaitophyta) sú fosílne dreviny z karbónu. Vyvinuli sa z prasličkorastov a významne sa podieľali na tvorbe ložísk čierneho uhlia.
Súčasné skupiny (triedy)
- Cykasorasty (Cycadophyta) sú „žijúce fosílie“, ktoré v druhohorách tvorili obrovskú časť vegetácie. Boli a sú to stromovité formy s hrubým, často šupinatým kmeňom a dlhými perovito zloženými listami na jeho vrchole. Hoci vizuálne pripomínajú palmy, patria k najstarším semenným rastlinám. Vytvárajú semená s nápadným mäsitým obalom (sarkotestou), ktoré vzhľadom pripomínajú kôstkovicu. Do súčasnosti sa zachovala len trieda cykasy (Cycadopsida). Sú striktne dvojdomé a na rozdiel od iných nahosemenných rastlín ich opeľuje hmyz (najmä chrobáky).
- Ginkorasty (Ginkgophyta) predstavujú jedinečnú a starobylú vývojovú líniu. Sú to druhotne hrubnúce stromy, ktoré najbohatšie zastúpenie dosiahli v triase a jure. Dnes prežíva už len jediný reliktný druh z triedy ginká (Ginkgopsida) a čeľade ginkovité (Ginkgoaceae) - ginko dvojlaločné (Ginkgo biloba). Je to dvojdomá „žijúca fosília“, nezmenená asi 150 miliónov rokov. Má charakteristické vejárovité listy s rovnobežnou žilnatinou, ktoré na jeseň opadávajú. Netvorí klasické šišky, ale mäsité semená s tvrdou kôstkou (pripomínajúce plody), ktoré po dozretí silne zapáchajú. Ginko sa celosvetovo pestuje ako veľmi odolná drevina.
- Borovicorasty (Pinophyta) tvoria najpočetnejšiu a evolučne najúspešnejšiu skupinu nahosemenných rastlín (asi 550 druhov).
- Lianovcorasty (Gnetophyta) sú evolučne najpokročilejšou skupinou a dôležitým prechodom ku krytosemenným rastlinám. Ako jediné nahosemenné rastliny už majú v dreve pravé cievy a u niektorých dochádza k dvojitému oplodneniu. Extrémnym a fascinujúcim zástupcom je velvíčia podivná (Welwitschia mirabilis). Je to reliktná rastlina z africkej púšte Namíb.

Ihličnany (Pinopsida)
Ihličnany sú evolučne veľmi staré dreviny, známe už od konca karbónu, anatomicky výborne prispôsobené extrémnym podmienkam. Majú úzke ihlicovité alebo šupinovité listy s hlboko vnorenými prieduchmi a hrubou kutikulou, čo ich výborne chráni pred mrazom a vyschnutím. Zvyčajne ide o jednodomé rastliny tvoriace drevnaté šišky. Sú však extrémne citlivé na znečistenie prostredia a kyslé dažde.
Čeľaď borovicovité (Pinaceae)
Majú vo svojich pletivách (kôra, drevo, listy) vytvorené živicové kanáliky, ktoré produkciou živice chránia rastlinu pred patogénmi a bylinožravcami. Rastú prevažne v severnom miernom pásme. Borovice sa od jedlí a smrekov odlišujú najmä tým, že ich ihlice vyrastajú zo skrátených konárikov (brachyblastov) vo zväzočkoch po dvoch, troch alebo piatich.
- Borovica lesná (Pinus sylvestris) - najrozšírenejšia domáca borovica.
- Borovica čierna (Pinus nigra) - pomerne často zdomácnelý druh.
- Borovica limbová (Pinus cembra) - rastie vo veľkých nadmorských výškach.
- Borovica horská (kosodrevina, Pinus mugo) - rastie nad hornou hranicou lesa.
- Jedľa biela (Abies alba) - dorastá do 40 m, má 3 cm dlhé ihlice so zárezom na špičke a belavými pásikmi na spodnej strane. Šišky rastú vzpriamene.
- Smrek obyčajný (Picea abies) - dosahuje výšku až 60 m. Ihlice sú štíhle, pevné a ostro zakončené. Šišky sú hnedé a visiace.
- Smrekovec opadavý (Larix decidua) - náš jediný opadavý ihličnatý strom.
- Duglaska tisolistá (Pseudotsuga menziesii) - často pestovaný nepôvodný druh.
Čeľaď cyprusovité (Cupressaceae)
Vždyzelené stromy a kry rozšírené po celom svete. Ich listy sú v dospelosti najčastejšie šupinovité (rastúce protistojne alebo v praslenoch).
- Borievka obyčajná (Juniperus communis) - má pichľavé ihlice v praslenoch. Vytvára čiernu, dužinatú a lesklú šišku (galbulus) pripomínajúcu bobuľu, ktorá sa vďaka vysokému obsahu silíc využíva v potravinárstve.
- Tuja západná (Thuja occidentalis) - častá severoamerická parková drevina s drobnými šupinovitými listami.
Čeľaď tisovité (Taxaceae)
Zahŕňa dvojdomé dreviny, ktorým chýbajú živicové kanáliky v dreve.
- Tis obyčajný (Taxus baccata) - dosahuje výšku 20 m, má ploché ihlice rastúce v dvoch radoch. Semeno chráni červený sladký miešok.
Čeľaď tisovcovité (Taxodiaceae)
Zahŕňa reliktné stromy pochádzajúce prevažne zo Severnej Ameriky a Ázie.
- Sekvoja vždyzelená (Sequoia sempervirens) - absolútny výškový rekordér, dorastajúca až do výšky 100 m.

Skrytý život stromov - Oficiálny trailer k filmu (2021)
tags: #rozmnozovanie #nahosemennych #rastlin